This article provides a basic overview of the immune system and how its successful engagement is necessary to produce a commercial vaccine, with a specific focus on the SARS-CoV-2 virus that causes the COVID-19 disease and the current pandemic. It also generally describes the different types of vaccines that could or are being used to make a commercial SARS-CoV-2 vaccine and identifies their current clinical status.
I. THE SMALLPOX VIRUS - A HISTORICAL EXAMPLE OF A VACCINATION
More than 200 years ago, the disfiguring and deadly smallpox virus was a major health issue until one scientist fully appreciated the significance of the phrase, “as smooth as a milk maid’s skin.” But how does that vain expression relate to vaccines? It connects the highly contagious causing smallpox virus with the much milder and related cowpox virus. In 1796, England’s Edward Jenner hypothesized why milkmaids, who routinely became infected with the cowpox virus, seemed to be immune to the smallpox virus and its disfiguring scars. He tested his hypothesis by inoculating a young boy with material harvested from a cowpox pustule, after which, the boy recovered from a mild cowpox illness. Two months later, Jenner inoculated the same boy with material from a smallpox pustule. Fortunately for the boy, he did not develop smallpox and he remained unscarred. (Reidel 2005.)
The scientific name for the cowpox virus is Variolae Vaccinae; its name partially derives from the Latin word for cow “vacca.” Thus, Jenner called this procedure “vaccination” and the material used in this process was called a “vaccine.” He devoted his life to promote vaccination and its use spread throughout England and Europe during his lifetime. In 1977, almost two centuries later, smallpox was eradicated as a result of vaccinations for that disease, which is a testament to Jenner’s life’s work. (Reidel 2005.)
The cowpox example demonstrates the basic principle of vaccination against a pathogen, typically a bacteria or virus that causes a disease. Any component in a vaccine that causes immunity against the pathogen is known as an “antigen.” An antigen is a molecule or a portion of a molecule recognized by the immune system as “foreign” that stimulates an immune response, which sometimes provides protective immunity from future infections. The cowpox virus vaccine induced protective immunity against the smallpox virus because the cowpox and smallpox viruses have structurally related antigens. In the case of the boy in Jenner’s experiment, his primed and prepared immune system was able to recognize and neutralize the incoming smallpox virus before it could cause disease. Thus, Jenner demonstrated that vaccination can protect an individual from disease.
II. THE IMMUNE SYSTEM – PROTECTORS OF THE BODY
The immune system is composed of an elaborate mixture of different cell types and cell signals that collaborate to provide a regulated and timely response to pathogens. The immune system can be sub-divided into two systems: (1) the innate immune system and (2) the adaptive immune system. The innate immune system is the body’s first line of defense that acts immediately or within hours of a pathogen’s appearance in the body. This system is a “generic” defense system in that it is not tailored to any specific pathogen. Conversely, the adaptive immune response develops over time to produce a tailored response that specifically targets the pathogen and includes antibody molecules and cells trained to recognize pathogens and pathogen-infected cells.
A. The Innate Immune System
- Physical Barriers
The innate immune system has multiple layers, but begins with physical barriers, such as the skin and mucus. The skin creates an unfavorable environment for pathogens, for example, because it is relatively dry, contains beneficial microbes as well as naturally occurring antimicrobial compounds against pathogenic microbes. Moreover, the skin’s outer layer is continuously sloughed off. Beneficial microorganisms on the skin play a symbiotic role by outcompeting pathogenic microbes. These microbes are present all over the skin as well as throughout the digestive tract. Mucus is another physical barrier that contains antimicrobial components, but it is also able to physically trap infectious agents, and in the respiratory tract, pathogens entrapped by mucus can be expelled by a productive cough. (Murphy et al. 2012.)
- White Blood Cells and Inflammation
White blood cells, which are important facilitators of inflammation, are another part of the innate immune system. Inflammation is a primary signal of, and is one of the first responses to, an infection. During inflammation, chemical signals called cytokines are released by damaged or infected cells. The cytokines are a diverse group of chemical compounds that convey instructions to cells surrounding the damaged/infected cells and are responsible for many of the physiological responses to an infection, such as dilation of blood vessels, heat and soreness. Cytokines signal the recruitment of other immune related white blood cells and stimulate defense mechanisms against intruders. For example, some white blood cells are equipped to detect the presence of unique viral RNA sequences and upon detection they release cytokines such as interferon, which activates a defense system that can capture the nascent virus. (Sparrer and Gack 2015; McNatt 2013 (discussing Tetherin protein capture of viruses).)
White blood cells are composed of a multitude of different cell types, are localized throughout the body, and are on constant surveillance for pathogens. Examples are:
- “Neutrophil” cells that neutralize a pathogen by releasing toxins,
- “Natural Killer” cells that kill infected host cells, thereby stopping pathogen reproduction in those cells,
- “Phagocyte” cells, meaning “eating cells,” that recognize, ingest and digest pathogens.
One kind of phagocytic cell, known as a dendritic cell, present pieces of the digested pathogen as antigens on its cell surface, which is then available to activate other immune cells. As explained below, these presented antigens help activate the adaptive immune system by “teaching” the cells involved in the adaptive immune system how to recognize and react to the pathogen. (Murphy et al. 2012.) This interplay between the two immune systems is discussed more in Section II. B. below.
- The Complement System
The complement system forms another part of the innate immune system. The complement system completes or “complements” the adaptive immune response because it comprises a set of specialized proteins in the blood that interact with “antibodies” (proteins produced by the adaptive immune system) and phagocytic cells to help clear foreign and damaged material. In a process called “opsonization,” antibodies and/or complement proteins coat a pathogen to facilitate its engulfment by phagocytic cells. In particular, complement proteins can bind with unique cell surface markers on the pathogen or with antibodies that have attached to the surface of a pathogen. These pathogen-bound complement proteins then interact with components on the cell surface of a phagocytic cell, which then engulfs and kills the pathogen. In addition, some complement components bind uniquely foreign pathogenic surface molecules, such as carbohydrates (made of sugars), and then destroy the pathogen by making holes in it. Finally, once the complement system has been activated, it causes interacting cells to release cytokines (i.e., chemical signals) that promote inflammation and recruit phagocytic cells. (Murphy et al. 2012.)
B. The Adaptive Immune System
The adaptive immune system develops over-time, potentially taking many days or weeks to develop. That is time well spent because it provides a tailored and specific response to the pathogen that can last months, years, or even a life-time. Its capacity to “remember” past pathogens prepares the immune system to immediately pre-empt a new infection from the same or a related pathogen. This quick and effective response was demonstrated by the cowpox/smallpox example discussed in Section I and is fundamental to successful vaccination.
- B-Cells and T-Cells
Cells that make up the adaptive immune system can be divided into two classes: B-cells and T-cells, each of which supply “specialized soldiers” to fight the pathogen. Each of these cell types originate from the bone marrow, but they mature in different places: B-cells remain in the bone marrow, whereas T-cells migrate to the thymus. Thus, their names signal the organ where they matured (“B” for bone marrow and “T” for thymus). During maturation, billions of B-cells and T-cells, each of which tailors its own unique cell-surface receptor, are selected or “trained” so that none of their receptors recognize “self components” in the body so as to eliminate any chance of autoimmunity (i.e., “friendly fire.”) The cell-surface receptor is like a “lock” that will recognize a specific “key,” which for present purposes is a pathogen-associated antigen. After maturation training, these newly formed naïve B-cells and naïve T-cells — or “naïve trained soldiers” — march via the bloodstream and lymphatic system and take residence in their “barracks,” known as the peripheral lymphoid organs, such as the spleen and lymph nodes. The naïve trained soldiers are considered “naïve” because they have not yet received their “orders” to be “activated” and unleashed against the enemy pathogen. (Murphy et al. 2012.)
- Development of the Tailored and Specific Immune Response
During an infection, information from the innate immune system gained from engaging with the pathogen is used to activate the adaptive immune system to mount a specialized and tailored attack on the enemy pathogen. Dendritic cells from the innate immune system perform “reconnaissance” and they follow the cytokine “flare” signals sent out by cells near the pathogen, which is where the battle is taking place. They are the immune system’s “super scouts.” They are “super” because they directly engage the pathogenic enemy and also pass on enemy information to the “specialized soldiers” (naïve B-cells and T-cells) of the adaptive immune system awaiting in their barracks. At the battle site, the dendritic super scout cells engage the enemy by digesting them and then displaying antigenic pieces of the enemy on their cell surface as “keys.” These cells are now “super scouts” that march to the “barracks” (e.g., the lymph nodes) with their antigenic keys proudly presented to the resident naïve B-cells and T-cells. T-cells lose their naïveté if their “locks” fit the antigenic “keys” presented by the dendritic cells, transforming them into “super trained soldiers,” able to identify enemy invaders to fend off the pathogenic enemy. (Murphy et al. 2012.)
The dendritic cell “super scouts,” and “super trained soldier” T-cells collaborate to present the pathogenic “key” to those “naïve soldier” B-cells that have the corresponding “key hole” in their “lock,” thereby converting those cells into “super trained” B-cells. Both classes of “super trained soldier” B-cells and T-cells, use the lock and key training method as a “learned visual” of the enemy in order to “seek and destroy” the pathogen. Thus, this training and selection by the adaptive immune system creates a tailored and specific immune response.
- Cell Number Expansion
Of the billions of potential soldiers, only a tiny fraction of them — the few and the proud — may have been activated with the “learned visual” antigens. Those few recruits, however, do not provide enough “troop strength” to defeat the enemy pathogen, which typically reproduce quickly, and so the number of recruits must be vastly increased. Consequently, both B-cell and T-cell “super trained soldiers” receive additional signals to clone themselves. By expanding the number of cells, the barracks begin to swell during an infection and that is why after infection the lymph nodes become swollen and tender to the touch. (Murphy et al. 2012.) (Note: do not touch them. Rather, let the soldiers expand and prepare for battle.)
- Specialized Capabilities
The newly cloned troops are also strengthened by receiving special fire-power capabilities, such as antibodies for “tagging and bagging” the enemy for destruction. Some of the super trained T-cells are transformed into one of three different kinds of specialized T-cell. The first kind become like “special forces” that gain the ability to recognize and kill infected cells. The second kind become like “drill sergeants” that help train and activate the immune system, such as the B-cells, as described above, to facilitate antibody production. The third kind become like “officers” who manage/regulate the battles/immune responses.
The activated and clonally expanded B-cells also have their weapon capabilities upgraded. Instead of just expressing the receptor “lock” on their cell surface, they are equipped to produce and release large amounts of free-floating clonal copies of the “locks” called “antibodies.” These free-floating antibodies are specialized and tailored weapons, like missiles, that home-in on the antigen “keys” on the enemy pathogen and with help from the complement system, tag them for death by immune cells (as discussed above in Section II.A.3). The antibodies can also independently neutralize the enemy. For example, in the case of a virus, after antibodies have covered all of the viral “keys,” the virus will be unable to use its keys to access the cellular locks to gain entry into the host’s cells, thereby blocking any further viral infection.
- Immunological Memory Cells
After the battle has been won, these specialized “super trained soldiers” cells and associated antibodies slowly wane. Yet, while some battles are forgotten as time passes, others are always remembered. Thus, often a sub-population of the trained and converted B-cell and T-cell “super trained soldiers” become “memory cells” after they retire from battle. These cells retain the memory of their specific enemy, but they will be re-activated as soon as their former pathogenic enemy (or a close resemblance) appears. Upon re-activation, these already primed and prepared cells rapidly arrive for battle to fight in the first response of any reinfection. This time around, they quickly expand their clonal troops and produce their specialized weapons, antibodies and special forces, to quash the enemy. Sometimes there is extra support already roaming the body — some antibodies from the last battle may still be present that can bind and neutralize the pathogen. This fast action alters the course of an infection to prevent disease. This is the principal purpose and role of vaccination – victory without war. (Murphy et al. 2012.)
III. THE IMMUNE SYSTEM AND VACCINATIONS
After it is administered, the ideal vaccine safely replicates the same immunological outcome from a natural infection (i.e., protective immunity), without causing disease or any substantial adverse effects. The adaptive immune response is the key to a successful vaccine. After vaccination, both memory B-cells and memory T-cells will be convinced that there had been a previous infection by the target pathogen that is now “remembered,” even though there was no prior battle. In essence, they will be “on guard,” surveying for what are considered to be “past foes.” T-cells derived from the memory T-cells will now be on hand so that they are ready to kill the pathogen and any newly infected cells harboring the pathogen. The memory B-cells will now be ready to replicate and produce massive amounts of neutralizing antibodies. Therefore, a successful vaccination will provide protective immunity and the affected individual may never realize there had been any contact with the pathogen. (Murphy et al. 2012.)
A. Adjuvants
For safety reasons, many vaccines developed today are not living organisms or infectious viruses like the cowpox example – that is they do not cause a disease. Jenner’s cowpox pustule vaccine caused a mild disease and contained a cross-reactive pox antigen to the smallpox virus, but there were probably other components in that pustule that helped to activate a robust and lasting immune response. (Riedel 2005.) However, infection with a related, mild virus, is rarely an option as a vaccine for a disease. Rather, modern vaccines typically have a selected subset of antigenic fragments from the pathogen, sometimes just a single fragment. Because of this, many modern vaccines need help because the reduced number of antigenic components from the pathogen on their own may fail to elicit a robust adaptive immune response. Such a vaccine design needs helping agents to activate a robust immune response. These helping agents are called “adjuvants,” because they help produce a fuller immune response. No adjuvant was required for the Jenner cowpox vaccine because it contained live cowpox virus that presented multiple antigens and persisted long enough to stimulate a robust immune response. (Murphy et al. 2012.)
Adjuvants are compounds that can stimulate phagocytic dendritic cells, which, as discussed above, are “super scouts” that digest the pathogens into small antigenic pieces and then present them as “keys” to other immune cells to achieve a full adaptive immune response. Adjuvants include a diverse group of chemicals, believed to act by different mechanisms, including aluminum salts, oil-in-water emulsions, and modified toxins from other pathogens, such as the pertussis toxin. (Murphy et al. 2012.)
Today, development of new adjuvants is a very active area of research because a limited number of adjuvants have been approved for use in humans and because modern vaccines, due to their reduced number of antigens, have failed when using the traditional adjuvants. (Shi 2019.) Often, each adjuvant must be tested with each potentially new vaccine. Some new adjuvants currently under investigation are based on molecules that are naturally found at a site of infection. These new adjuvants include structures that are unique to pathogens, for example: (1) flagellin, a protein that helps bacteria move, and (2) double stranded viral RNA, a hallmark of RNA viruses, such as SARS-CoV-2, discussed below. In addition, cytokines involved in host immunological signaling are being investigated as new adjuvants. (Murphy et al. 2012; Shi 2019.) For example, in the development of vaccines against the virus that causes COVID-19, the pharmaceutical company GlaxoSmithKline (GSK) has decided to share its proprietary vaccine adjuvant named “AS03.” GSK has been collaborating with four vaccine developers, including Sanofi. (Walker 2020.) The AS03 vaccine adjuvant is an oil-in-water emulsion made from squalene (a small molecule that animals use to make cholesterol), vitamin-E (e.g., DL-α-tocopherol) and polyoxyethylene sorbitan monooleate (e.g., the detergents, polysorbate 80 or tween 80 that are added to some foods). (Walker 2020; US Patent 9,700,605; Wu 2019.)
IV. VIRUSES AND THE SARS-COV-2 VIRUS’S RELATIONSHIP TO COVID-19
A. Viruses
Viruses can be considered as tiny packets of genetic information. The amount of genetic information in a virus is minuscule compared to a human cell, consisting of as few as only thousands of nucleotides. In contrast, a human cell nucleus contains around 3 billion nucleotides. Viruses cannot replicate on their own, but rather they rely on a host’s cellular machinery in order to reproduce. In general, viruses encode just enough genetic information: (1) to produce the equipment necessary to hi-jack the host cell’s machinery for itself, (2) to avoid the host’s immune defenses, and (3) to produce unique viral components, such as viral coat proteins.
There are two classes of viruses: (1) enveloped viruses, which are encased in a lipid (i.e., fatty) membrane that contains viral proteins and (2) non-enveloped viruses, which have a more robust and durable outer layer (i.e., capsid shell) made of proteins. Both classes of virus contain “spike” proteins on their surface that act as “keys” to gain entry into a host cell. The spike protein “key” will only fit the right “lock,” which is a receptor on the host’s cell surface that the virus uses to locate a susceptible cell for infection. After the key-lock connection has been made, the virus gains control of the host cell and the infection begins. As described above, if antibodies block the viral “keys,” the virus cannot bind to the host cell and it is neutralized. A major role of vaccination is to have these protective neutralizing antibodies already present to stop an infection.
B. SARS CoV-2 Virus and the COVID-19 Disease
It is important to appreciate the distinction between a virus and the disease that it causes. The scientific name of the virus that has caused the present pandemic is “Severe “Acute “Respiratory “Syndrome “Corona“virus “2” (SARS CoV-2) or the ““2019 Novel “Corona“virus” (2019-nCoV). Only a sub-set of infected individuals develop the disease named “Corona“virus “Disease 20“19 (COVID-19) when infected by SARS CoV-2. This is conceptually like HIV-1 and AIDS, where the Human Immunodeficiency Virus 1 (HIV-1) is the virus that causes the disease called “Acquired Immune Deficiency Syndrome (AIDS).” Not everyone who is infected with HIV-1 has or will develop AIDS, as is the case for SARS CoV-2 and COVID-19.
A vaccine should protect the site of primary infection. The SARS CoV-2 virus infects the epithelial (outer layer) cells in lungs and nasal passages and can spread to infect other less accessible blood vessel endothelial cells that line the interior surface of its lumen. (Hou 2020.) The SARS CoV-2 virus is an enveloped virus and its spike protein “key” recognizes the host human cell receptor “lock” protein, which is known as Angiotensin-converting Enzyme 2 (“ACE-2”). (Callaway 2020.) The human ACE-2 protein is found on various cell types, including: (1) those lining the nasal passage, (2) those lining the lung alveolar cells (these facilitate exchange of oxygen and carbon dioxide gases), (3) endothelial blood vessel cells found throughout the body, (4) arterial smooth muscle cells, and (5) enterocytes (cells of the intestinal lining). (Hamming 2004; Hou 2020.) Because they are exposed to air, alveolar and nasal cells are the most susceptible cells for infection from the SARS CoV-2 virus, which is spread through airborne microdroplets. (Hou 2020.) After entry into and replication in alveoli, the viruses can exit towards the blood vessels. Thus, after infection of the lung alveoli, it is possible the adjacent endothelial cells in the blood vessels become a secondary site of infection, which may be one way the infection spreads and becomes systemic. (García 2020; Nova 2019.) An ideal vaccine should provide protection to the alveolar and nasal epithelial cells of the respiratory tract to stop any potential secondary infections of the blood vessels.
The mutation rate of SARS CoV-2 is low, which bodes well for vaccine development. For more rapidly mutating viruses such as flu or HIV-1, successful vaccines have been either elusive, as for HIV-1 or have to be redeveloped each season, as for flu. (Callaway 2020.) The lack of success for HIV-1 and flu vaccines may be because the rapidly mutating viruses stay steps ahead of the immune system soldiers. So far, the mutations that have accumulated in the SARS-CoV-2 virus have not changed the capability of antibodies to neutralize the virus or provided an advantage for the virus. (Callaway 2020.) This may mean that additional development of vaccines will be unnecessary beyond the first round of successful vaccines currently being developed against SARS-CoV-2. See Table 1, below. However, how long an individual’s immunity lasts against that virus (see discussion above re: memory cells at Section II. B. 5) has yet to be determined, meaning that re-vaccination may be necessary.
V. DIFFERENT TYPES OF VACCINES
Various types of modern vaccines are generally described in this section, along with the pros and cons of each type. The various universities, institutes and pharmaceutical companies developing vaccines against SARS-CoV-2 are identified and classified according to the type of vaccine and its clinical stage in development in Table 1, below.
An obvious benefit of having multiple different vaccines in development is the increased chance of finding at least one successful vaccine. There are other benefits. Different vaccines may be useful because the virus could mutate to escape any one vaccine, making the “escaped” vaccine of little use. Additionally, different vaccines may provide protection against other related pathogenic viruses if those viruses are cross-reactive to one of the vaccines, as in the cowpox/smallpox example. In-fact, the “spike protein” from the related SARS-CoV from 2003 is 75% identical to the SARS CoV-2 “spike protein” and neutralizing antibodies from the SARS-CoV can also neutralize SARS-CoV-2. (Hou 2020.) Yet another reason for having different vaccines is that some may not work in certain human populations because not all individuals are able to present the same vaccine derived antigen for training the B-cells and T-cells. Finally, different vaccines can benefit vaccine production capacity if different manufacturing methods are used to make the different vaccines, as that can reduce competition for limited resources.
A. Live-attenuated Virus Vaccines
- General Description
Live attenuated virus vaccination requires an infectious virus that has been mutated to become substantially weakened (i.e., attenuated) so that it causes no more than mild symptoms. Also essential is that the attenuated virus have the required antigen(s). The cowpox vaccine is a good example of an attenuated virus vaccine, which caused a much milder disease but resulted in protective immunity from the harsher smallpox virus. Modern genetic engineering enables precise and controlled mutagenesis of many viruses to reduce their pathogenicity. Historically, testing on attenuated virus vaccines for safety and efficacy has occurred over a relatively long period before they have been approved. This has been the case for the live-attenuated virus vaccines that are currently used to vaccinate against polio, measles, mumps, rubella, and varicella. (Murphy 2012). Because the SARS-CoV-2 virus has only recently been discovered and because the need for a vaccine is urgent, there presently is no effort to develop a live-attenuated vaccine for SARS-CoV-2.
- Pros
Advantages for live-attenuated virus vaccines include (Murphy 2012):
- Increased efficacy since viruses reproduce and persist longer, which increases the opportunity for the immune system to select for strong antigens
- multiple antigens are introduced:
- this increases the chance that one or more of these antigens will provide protective immunity
- an immune response to multiple antigens decreases the chance the pathogen can mutate to avoid immunity
- typically does not require an adjuvant
- usually only one administration is necessary
- Cons
Disadvantages for live-attenuated virus vaccines are due to decreased safety. For example, these vaccines (Murphy 2012, Riedel 2005):
- may cause disease (the boy in Jenner’s experiment developed mild disease due to the cowpox vaccine)
- may be lethal when administered to immunocompromised individuals
- Such people must rely on either an alternate vaccine or on “herd immunity,” which is where a high percentage of individuals have immunity so that the disease transmission rate is essentially nonexistent
- the attenuated virus can revert to the original pathogen
- In rare cases, the gene mutation(s) used to attenuate the virus may change back to the original gene sequence
B. Inactivated Virus Vaccine and Killed Virus Vaccine
- General Description
Inactivated and killed virus vaccines are typically made by growing and harvesting virus-like particles or viruses, respectively, in a controlled environment, after which they are treated by chemical, heat, or radiation treatment. To increase safety, an inactivated virus vaccine uses virus-like particles that are genetically inactivated. These virus-like particles are bioengineered to alter key genes and proteins of the virus to render it non-infectious. For example, it is common to remove most of the genetic material from the virus, which makes virus-like particles that are gutted shells of their former native versions. (Mohsen 2017.) Examples of inactivated vaccines include hepatitis B, human papillomavirus, and malaria. (Mohsen 2017.) Examples of killed virus vaccines include polio, rabies, hepatitis A, cholera, plague and most influenza vaccines. (Murphy 2012.)
- Pros
Advantages for inactivated/killed virus vaccines include:
- increased safety over live-attenuated vaccines:
- inactivated viruses with gross or multiple mutations can block reversion back to the native virus
- properly killed viruses are non-infectious
- increased effectiveness since a number of viral antigens will be present
- Cons
Disadvantages for inactivated/killed virus vaccines include:
- may elicit weaker immune responses because:
- there is no chance for reproduction, which means viral antigens will not persist relative to a live virus
- may not invade cell and activate internal cellular responses
- for inactivated virus vaccines the number of antigens are reduced relative to live or killed virus vaccines
- incomplete inactivation or killing can result in live virus infection
- may require adjuvants
- booster shots may be required for desired immunity, which requires record keeping and suffers from reduced patient compliance
- resource crowding, vaccine harvesting may compete with flu vaccine production, which reduces scalability
C. DNA Vaccines
- General Description
DNA vaccines are composed of DNA fragments that encode viral proteins or protein fragments (i.e., peptides). The DNA encodes either surface or internal proteins of the virus. Internal viral proteins are chosen if they are predicted to be strongly antigenic and would be expected to be presented at the cell surface of infected cells, which would then be targeted by the immune system for killing. Surface viral proteins are often based on the “spike protein” that the virus uses to infect a host cell so that the vaccine induces antibodies that can neutralize the pathogen, as discussed in Section IV.
The DNA is then introduced to the host via an injection, usually in the deltoid muscle, and some small percentage of the DNA finds its way into host cells. (Liu 2019.) But to increase cellular entry of the DNA, there are generally two methods of delivering the DNA. The first method packages the DNA into nano-particles that facilitate entry into the cells. (Dalirfardouei 2020.) The second method does not package the DNA but rather relies on electroporation. Briefly, at the site of the injection, mild electric-pulses produce nano-holes in the cell surface. The nano-holes and electricity then facilitate the DNA to enter the cell. This electroporation method is used by Inovio Pharmaceuticals to deliver a potential DNA vaccine against COVID-19. See Table 1 below. (US Patent 7,328,064; US Published Patent Application No. 2019/0284263.)
The modified host cells use their machinery to make the mRNA corresponding to the vaccine DNA sequence, which directs the production of the viral protein/peptide. To the extent that the protein/peptide is made available to the immune system and is a good antigen, it will be recognized as foreign and elicit an immune response. As of 2018, there were no approved DNA vaccines for human use, although at that time there were more than 500 clinical trials on DNA vaccination. There is, however, one approved DNA vaccine for horses against the West Nile Virus. (Hobernik and Bros 2018.)
- Pros
Advantages of a DNA vaccine are its safety and production benefits (Hobernik and Bros 2018):
- no risk of pathogen infection
- quick production using machines for synthesis or via bio-reactors for scale-up
- cost effective
- very stable, storable at ambient temperature
- can provide multiple pathogenic antigens, which is useful for
- pathogens that mutate quickly
- increase a stronger immune response
- adjuvants can be encoded into the DNA (e.g., adding a DNA sequence that encodes a cytokine)
- extended production and thus lifespan in the host of the introduced protein(s)/peptide(s), which can increase efficacy beyond protein or inactivated/killed vaccines
- Cons
Disadvantages of DNA vaccines relate to vaccine composition (Murphy 2012; Hobernik and Bros 2018.):
- fewer viral antigens are presented to the immune system
- booster shots may be required for desired immunity, which requires record keeping and suffers from reduced patient compliance
- may require add-on adjuvants if the adjuvants are not encoded in a DNA fragment in the vaccine
- potential for integration into the host genome that can cause host mutation
D. mRNA Vaccines
- General Description
Conceptually, mRNA vaccines are very similar to DNA vaccines except: (1) the mRNA is directly encoded into protein/peptide and (2) the mRNA is typically packaged into lipid nano-particles, which protects the mRNA and facilitates delivery to the host cells, with or without electroporation.
- Pros
Advantages to an mRNA vaccine are similar to DNA vaccines, but there are also added benefits:
- integration into and mutagenesis of the genome is precluded
- production of the protein/peptides is faster than DNA vaccines.
- Cons
Disadvantages to mRNA vaccines are similar to DNA vaccines except that:
- mRNA vaccines are less stable
- must be stored in a controlled environment
- require special formulation (e.g., lipid nano-particles)
- shorter duration in the body than DNA vaccines:
- the mRNA may require unconventional or chemically modified bases in the RNA (for example pseudouridine) (Liu 2019)
- may produce less antigenic protein over a shorter time period than DNA vaccines, reducing protective immunity
- new method of vaccination, an mRNA vaccine has yet to be commercialized, although Moderna and Pfizer with BioNTech and Fosun Pharma are close to launching mRNA vaccines (presently in Phase III clinical trials, see Table 1, below).
E. Protein and Peptide Vaccines
- General Description
Protein vaccines are composed of antigenic proteins or peptides derived from the pathogen. These vaccines are typically made using bioengineering techniques where DNA that encodes the pathogenic protein or peptide is introduced into a bacteria, yeast, or a mammalian cell line that allows for growth in large scale bioreactors. The purified antigenic protein or peptide is then formulated into a vaccine and injected into the individual. This method has been successfully used to vaccinate against the hepatitis B virus. (Murphy 2012.)
- Pros
Advantages for protein/peptide vaccines are:
- safety because to date they have not caused any viral related disease
- production is scalable
- can meet high demand
- Cons
Disadvantages for protein/peptide vaccines are:
- production of the protein or protein fragment (peptide):
- can be laborious and costly
- may not be predictably scalable, depending on the particular protein or peptide
- stability issues may require special handling and storage
- smaller repertoire of protein/antigens:
- smaller immunological presentation may result in an inadequate immune response
- allows for greater chance for virus to mutate to escape the vaccine
- booster shots may be required for desired immunity:
- short lifespan of the protein or peptide in the body
- requires record keeping and suffers from reduced patient compliance
- often require an adjuvant
TABLE 1. ENTITIES ACTIVELY PURSUING A VACCINE FOR COVID-19 CURRENTLY IN CLINICAL EVALUATION
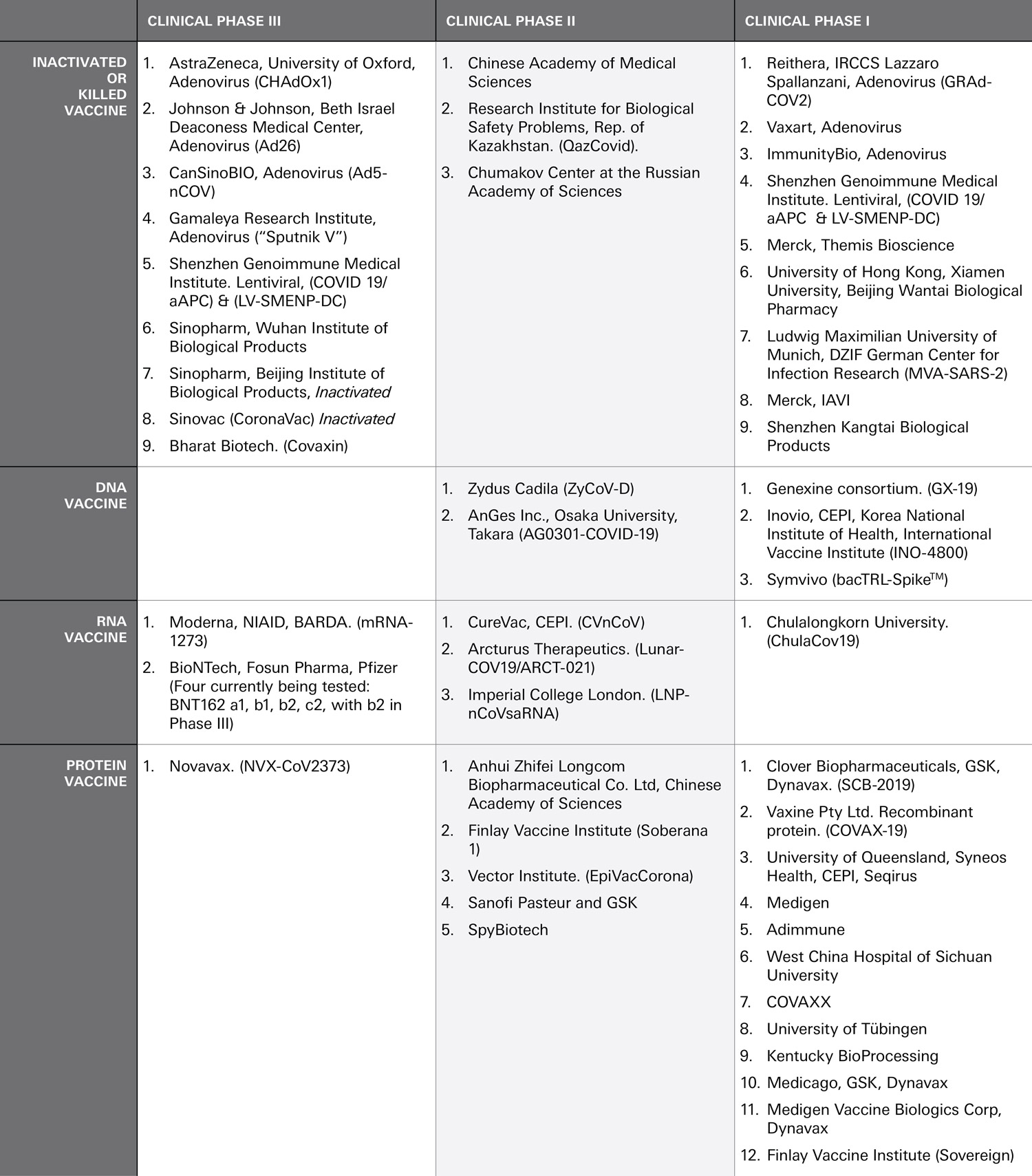
(Corum 2020; Neubert 2020.)
Name of vaccine being developed is provide in parenthesis ( ) after the company name.
All vaccines in combined phases were categorized into the further along higher phase.
REFERENCES
Riedel, Stefan. “Edward Jenner and the History of Smallpox and Vaccination.” Proc (Bayl Univ Med Cent) 18, no. 1 (January 2005): 21–25.
Murphy, Kenneth, Paul Travers, Mark Walport, and Charles Janeway. Janeway’s Immunobiology. New York: Garland Science. 2012.
Sparrer, Konstantin MJ, and Michaela U Gack. “Intracellular Detection of Viral Nucleic Acids.” Curr Opin Microbiol 26 (August 2015): 1–9.
McNatt, Matthew W., Trinity Zang, and Paul D. Bieniasz. “Vpu Binds Directly to Tetherin and Displaces It from Nascent Virions.” PLoS Pathog 9, no. 4 (2013): e1003299.
Shi, Shuting, Haoru Zhu, Xinyu Xia, Zhihui Liang, Xuehu Ma, and Bingbing Sun. “Vaccine Adjuvants: Understanding the Structure and Mechanism of Adjuvanticity.” Vaccine 37, no. 24 (05 27, 2019): 3167–78.
Walker, Joseph. “To Find a Coronavirus Vaccine, GlaxoSmithKline Is Bonding With Its Biggest Competitors.” The Wall Street Journal, October 24, 2020, sec. B, p1.
Wu, Biqing, Dieyckson O. Freire, and Richard W. Hartel. “The Effect of Overrun, Fat Destabilization, and Ice Cream Mix Viscosity on Entire Meltdown Behavior.” J Food Sci 84, no. 9 (September 2019): 2562–71.
Patent, William Ripley Ballou JR, Emmanuel Jules Hanon, Vaccine comprising an oil in water emulsion, registered as No. US9700605B2, January 9, 2015.
Hou, Yixuan J., Kenichi Okuda, Caitlin E. Edwards, David R. Martinez, Takanori Asakura, Kenneth H. Dinnon, Takafumi Kato, et al. “SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract.” Cell 182, no. 2 (07 23, 2020): 429-446.e14.
Callaway, Ewen. “The Coronavirus Is Mutating — Does It Matter?” no. 7824. Nature 585, no. 7824 (September 8, 2020): 174–77.
Hamming, I, W Timens, MLC Bulthuis, AT Lely, GJ Navis, and H van Goor. “Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis.” J Pathol 203, no. 2 (June 2004): 631–37.
García, Luis F. “Immune Response, Inflammation, and the Clinical Spectrum of COVID-19.” Front. Immunol. 11 (2020).
Nova, Zuzana, Henrieta Skovierova, and Andrea Calkovska. “Alveolar-Capillary Membrane-Related Pulmonary Cells as a Target in Endotoxin-Induced Acute Lung Injury.” no. 4. International Journal of Molecular Sciences 20, no. 4 (January 2019): 831.
Mohsen, Mona O., Lisha Zha, Gustavo Cabral-Miranda, and Martin F. Bachmann. “Major Findings and Recent Advances in Virus-like Particle (VLP)-Based Vaccines.” Semin Immunol 34 (12 2017): 123–32.
Liu, Margaret A. “A Comparison of Plasmid DNA and MRNA as Vaccine Technologies.” Vaccines (Basel) 7, no. 2 (April 24, 2019).
Dalirfardouei, Razieh, Mohsen Tafaghodi, Zahra Meshkat, Adel Najafi, Aida Gholoobi, Maryam Sadat Nabavinia, Samineh Sajedifar, et al. “A Novel Formulation of Mtb72F DNA Vaccine for Immunization against Tuberculosis.” Iran J Basic Med Sci 23, no. 6 (June 2020): 826–32.
Patent, Iacob Mathiesen, Torunn Tjelle, Knut Arvid Sørensen Rekdahl, Bjøm David-Andersen, Electroporation device and injection apparatus, registered as No. US7328064B2, July 3, 2003.
Patent Application 2019/0284263, Trevor Smith, Nina Schommer, Kate Broderick, Bryan Yung, Katherine Schultheis, In Vivo use of Chondroitinase and/or Hyaluronidase to Enhance Delivery of an Agent, filed as No. 16/302,567.
Hobernik, Dominika, and Matthias Bros. “DNA Vaccines—How Far From Clinical Use?” Int J Mol Sci 19, no. 11 (November 15, 2018).
Corum, Jonathan, Sui-Lee Wee, and Carl Zimmer. “Coronavirus Vaccine Tracker.” The New York Times, November 3, 2020, sec. Science. https://www.nytimes.com/interactive/2020/science/coronavirus-vaccine-tracker.html (Last Accessed November 6, 2020)
Neubert, Karola, Janna Schmidt, and Anja Brandt. “First Clinical Trial of DZIF Vaccine against COVID-19 Gets Go-Ahead.” DZIF German Center for Infection Research, Press Release October 2, 2020, https://www.dzif.de/en/first-clinical-trial-dzif-vaccine-against-covid-19-gets-go-ahead (Last accessed November 6, 2020)